No new proteins, no loss of proteins, just changes in gene expression levels.
The hallmark of evolution is repurposing, hardly consistent with any omnipotent designer. Therefore, there’s zero biological basis for @Giltil’s applying anything in that PLoS paper to vertebrate limb differences.
It’s completely irrelevant. Those who have been taken in by the ID scam generally don’t bother to see how irrelevant it is to how biology is known to work. In your case, you are assuming that new proteins must be involved in changing limb morphology, on the basis of absolutely nothing but wishful, simplistic thinking.
Given your continuous output of gross falsehood on the forum (gold medal performance here), you are not entitled to my time to “tell you more.” The answer to your question is easily found in the article you were quotemining. Instead of “telling you more” I will ask you a few basic questions. If you answer a couple of them, you might earn an explanation.
Did you read the paper you cited? Specifically, did you read the Discussion?
Have you read any recent paper on fitness landscapes? I mentioned two in this very thread.
Do you know what recombination is and why it is relevant in the 18-year-old PLOS ONE paper that you posted while ignoring the two papers I just mentioned?
Do you read scientific papers before posting on the forum? For example, did you even check to see if the what you typed here is even close to accurate?
First one would need to ask the question of whether such a modification is even evolutionarily possible?
AFAIK, no (viable) member of superclass Tetrapoda, or even its parent clade Teleostomi, has produced even a rudimentary third pair of limbs.
Where would this third pair of limbs even attach to the skeleton? What genetic modifications would be necessary for even a rudimentary third pair? Does a neutral/adaptive pathway to these modifications even exist? These are the sorts of question that need to be delved into before asking “why evolution neither produced animals having both of these structures” becomes a valid question.
Lacking such answers, this scenario appears to be just another example of the empty rhetoric that typifies ID apologetics, be it from Gil, Miller, Dembski, Behe, Meyer, or the rest of the DI crew.
If ever a candidate existed for something that is genuinely beyond the “edge of evolution”, this scenario would appear to be it.
It seems to me that @Giltil engaged in quote mining because he could not answer the actual question. He could not produce any barrier to limb evolution so he removed all that in the hope that we would think that any limit on RV + NS would do. But given the fuller context the paper is obviously irrelevant to the question actually asked.
What’s worse, the paper doesn’t even support his beliefs about protein evolution. He seems to get tunnelvision on the 1070 number and everything else the paper says on that same topic fails to register in his mind.
No. I nonetheless wanted to read the first one you mentioned, but unfortunately it’s access is subject to a fee. Now, I note in the abstract that the authors describe a rugged fitness landscapes but that this rugged landscape has multiple properties of a smooth landscape, which certainly facilitates adaptive evolution. But not all rugged fitness landscapes have these facilitating properties, have they?
Yes, I know what recombination is but I disagree with you and the authors that it has relevance here.
Well, no, I didn’t check. Unfortunately, for if I had, I would have seen that I was wrong. Sorry for that.
How sad that you don’t have access to a library (or an employer) that subscribes to Science. My condolences. Go to Wagner’s website, navigate to the paper (“A rugged yet easily navigable fitness landscape of antibiotic resistance”), click on “reprint request.” This takes you straight to the PDF.
That’s really not accurate. A rugged landscape is just rugged; if you zoom in to any one part of any large rugged landscape, you will find areas with “properties of a smooth landscape.” That’s not what the paper shows. It shows that there are adaptive paths through the rugged landscape. And perhaps more importantly, it shows that the ruggedness of a landscape does not (as is asserted by shallow misinformation-creating writers at anti-evolution propaganda organizations, including the ignorant writer you cited to begin this thread) inherently result in vast barriers to adaptation.
Certainly not. Wagner and colleagues are explicit about this. But also, the second paper I mentioned in this thread explores how extremely rugged landscapes can act as (this is my word) “locks” on adaptive choices, so that specific interactions can stay that way. You can read a preprint of that paper here or ask me to provide the PDF.
This means that simplistic religious-like “rules” don’t apply. That’s true of essentially all of biology, and you should have learned it a long time ago.
This means that you don’t know what recombination is, or that you did not understand the paper at all, or that you are doing what you do too often on this forum, which is type BS without bothering to even check on whether your typing matches reality. You actually can’t “disagree” that recombination “has relevance here,” just like you can’t “disagree” about continental drift or human genetic diversity. And personally speaking, I find this disappointing, @Giltil. Because you just did something good that ID people almost never do:
Here you admitted error, and even apologized for it. You admit not only that you were scientifically wrong but that you were wrong to write something without checking to see if it was correct. Thank you for doing that! It’s an essential intellectual practice but it’s also a way to show respect to others in a discussion. That’s why I’m disappointed in your claim to “disagree” with a statement of plain clear undisputable fact about recombination in a phage experiment. You can’t make that statement with any intellectual credibility, and by doing it anyway, you show (unintentionally, I am sure) disrespect for people in the forum. Bill Cole does this reflexively and repeatedly. I’d personally love to see you do better than him.
Right, and penguins more or less “fly” underwater. It might be harder to evolve into a brachiating mode of movement, especially if there are monkeys already in that niche.
How do you know this? What are the evidence for this claim that seems completely unsubstantiated to me given, for example, the very low level of sequence homology between the two domain?
Both sequence-similarity in key regions, but also secondary and tertiary structure similarities. See(links to pdf of these papers can be found on google scholar):
Holliger P, Riechmann L, Williams RL. Crystal structure of the two N-terminal domains of g3p from filamentous phage fd at 1.9 A: evidence for conformational lability. J Mol Biol . 1999;288(4):649-657. doi:10.1006/jmbi.1999.2720
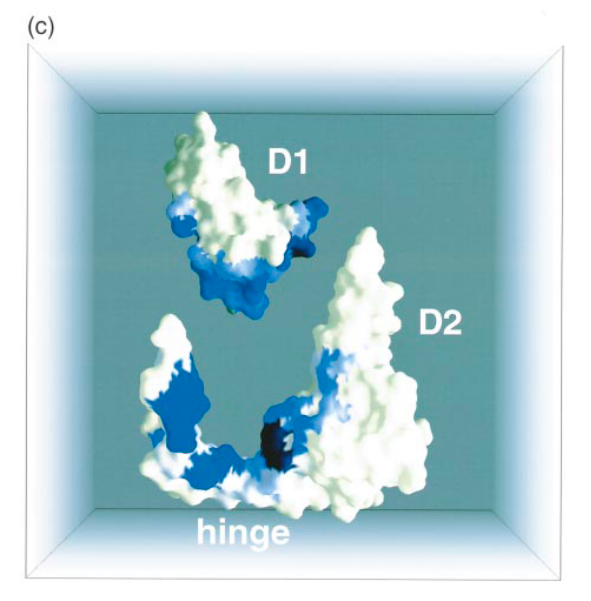
Several features make the hinge sub-domain (HD) stand out from the rest of the structure. It has been proposed that the core of D2 (residues 124-202; D2 (Figure 1(a)) may have arisen from D1 by gene-duplication (Lubkowski et al., 1997) and indeed, both D1 and D2 are compact b-barrel domains of identical topology with a short a-helix packing onto the outside of the barrel. In contrast, the hinge sub-domain seems unlikely to have arisen as part of this gene duplication. Rather, D2* appears to be an insertion into the previously continuous sequence of the HD. The fact that hinge sub-domain makes the majority of its contacts with D1 (12 compared to 6 with D2*, Figure 1(a) and (c)) also suggests that it may have predated the gene-duplication that gave rise to D2*. Intriguingly, most of the buried contacts between HD and D1 (Figure 1(a) and (c)) involve residues predicted by NMR to be part of the TolA-binding site on D1 (Riechman & Holliger, 1997).
Lubkowski J, Hennecke F, Plückthun A, Wlodawer A. The structural basis of phage display elucidated by the crystal structure of the N-terminal domains of g3p. Nat Struct Biol . 1998;5(2):140-147. doi:10.1038/nsb0298-140
Especially in the 2nd paper they show the similarity in the order of secondary structural elements, with key residues also being similar along the structure, and a nice picture of the tertiary structures from the D1(blue) and D2(red) domains superimposed on each other:
Notice the order and direction of the beta sheets β8, β9, β10 and β11 in the D2 domain are in the same order and direction as the sheets β2, β3, β4 and β5 in the D1 domain. Also the alpha helix α2 that comes before β12, and α1 before β1:
Figure 1. (a) Sequence of the D1D2-fd. The sequence is overlaid with secondary structure elements. Rounded boxes and arrows represent a-helices and b-sheets, respectively. Residues visible in the electron density map are in bold print. The hinge domain region is shown in italic print and underlined by a broken line. Hinge residues G99 and S208 are highlighted. Circles underneath residues indicate that residues are buried in the D1D2 structure but not in isolated domains. Shading reflects degree of burial of residues in the D1, D2 interface (see (c)), white reflects the lowest degree, grey an intermediate and black the highest degree of burial. Domains D1 and D2* are shown within square brackets. (b) Ribbon representation of D1D2 fragment from phage fd (D1D2-fd): domain D1 (2-66) is coloured yellow, the homologous b-barrel core of D2 (D2*) is coloured green , and the proposed hinge sub-domain (HD) including strands b6 and b13, is coloured cyan. Secondary structure elements are labelled throughout. Hinge residues G99 and S208 are shown as spheres. (c) Space-filling representation of D1D2-fd. D1 (2-66) is moved away from the rest of the molecule. The colour coding for residues represents the degree of burial as measured by the difference in solvent accessibility (using a probe sphere of 1.4 A) in the individual D1 and D2 domains and in the D1D2-fd. Degree of burial varies fromwhite (fully exposed) over blue to black (completely buried).
Several features make the hinge sub-domain (HD) stand out from the rest of the structure. It has
been proposed that the core of D2 (residues 124-202; D2* (Figure 1(a)) may have arisen from D1 by gene-duplication (Lubkowski et al., 1997) and indeed, both D1 and D2* are compact b-barrel domains of identical topology with a short a-helix packing onto the outside of the barrel. In contrast, the hinge sub-domain seems unlikely to have arisen as part of this gene duplication. Rather, D2* appears to be an insertion into the previously continuous sequence of the HD. The fact that hinge sub-domain makes the majority of its contacts with D1 (12 compared to 6 with D2*, Figure 1(a) and (c)) also suggests that it may have predated the gene-duplication that gave rise to D2*. Intriguingly, most of the buried contacts between HD and D1 (Figure 1(a) and (c)) involve residues predicted by NMR to be part of the TolA-binding site on D1 (Riechman & Holliger, 1997). This raises the possibility that the HD is involved in giving some conformational Flexibility to the D2 domain and is a major component of the blockage of the TolA-binding site on D1
Here is what the authors of the paper you referred to say: It has been proposed that the core of D2 (residues 124-202; D2 (Figure 1(a)) may have arisen from D1 by gene-duplication (Lubkowski et al., 1997)
Here is what you said: Incidentally the D2 domain of the g3p protein evolved by an internal duplication and divergence of another domain in the same protein
In the one case, a plausible but highly hypothetical scenario is offered, in the other case, said scenario is erroneously elevated to the level of a fact.
Highly hypothetical? What else do diverged duplicates look like?
No, sorry. This is a fact. This is a diverged duplicate, and structual and sequence similarity analysis is how we recognize that to be a fact. It happened. Not hypothetically. This is what it would look like if it happened.
How could it progress to fact for you, if not in this way? Must we have been there?
No, it is not a fact. If this was the case, why didn’t the authors recognize it as such. Why did they say « It has been proposed that the core of D2 may have arisen from D1 by gene-duplication » instead of something like « we know that the core of D2 have arisen from D1 by gene-duplication »? Note that if they had said the latter, they wouldn’t have passed the peer reviewed process. And for good reason.
You are here guilty of « affirming the consequent», a logical fallacy.
Among other reasons beacuse different people have different thresholds for what should be considered a fact (and there isn’t a sort of objective measure of it), but in this case probably more importantly because scientists generally write in this way where they kinda hedge their statements.
Pretty much the entirety of the field of biology considers evolution a fact, and that’s written in countless papers and textbooks. But you don’t agree, so who cares whether the paper said it anyway?
We have no idea whether it would have. It’s pointless speculation.
You seem to have confused proof with fact. I’m not making a deductive argument, or doing math. It’s an inference to the best explanation. I can’t prove there’s milk in my fridge, it will always remain an inference we draw from observations.
Inferering a gene duplication from structural and sequence similarity really isn’t that much more complicated than finding footprints in dried mud and inferring someone walked there barefeet when it was wet. It would be ridiculous to deny it.
But hey, if you don’t want to call it a fact, then fine. Not clear what you think it takes to be a fact, you’ve avoided answering my question. But you said it was plausible, and that’s good enough for you to not have a case here. Either way the paper can’t support any of the statements made by you or the Evolution News article you linked in the op