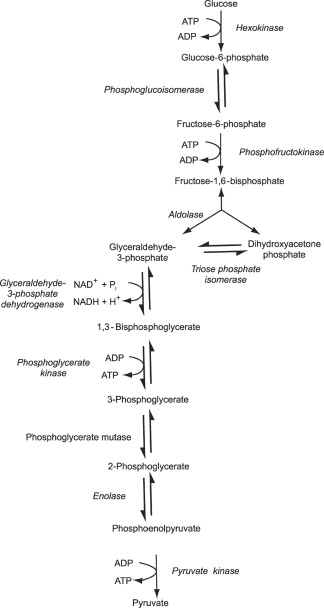
Cards lying flat on the ground or some flat surface is a highly probable configuration from random orientations and positions and velocities over time for the cards, etc.

In contrast, the following is not a highly probable configuration from random orientations and positions and velocities over time for the cards:
The house of cards is improbable in the statistical sense of the space of trajectories that can construct it.
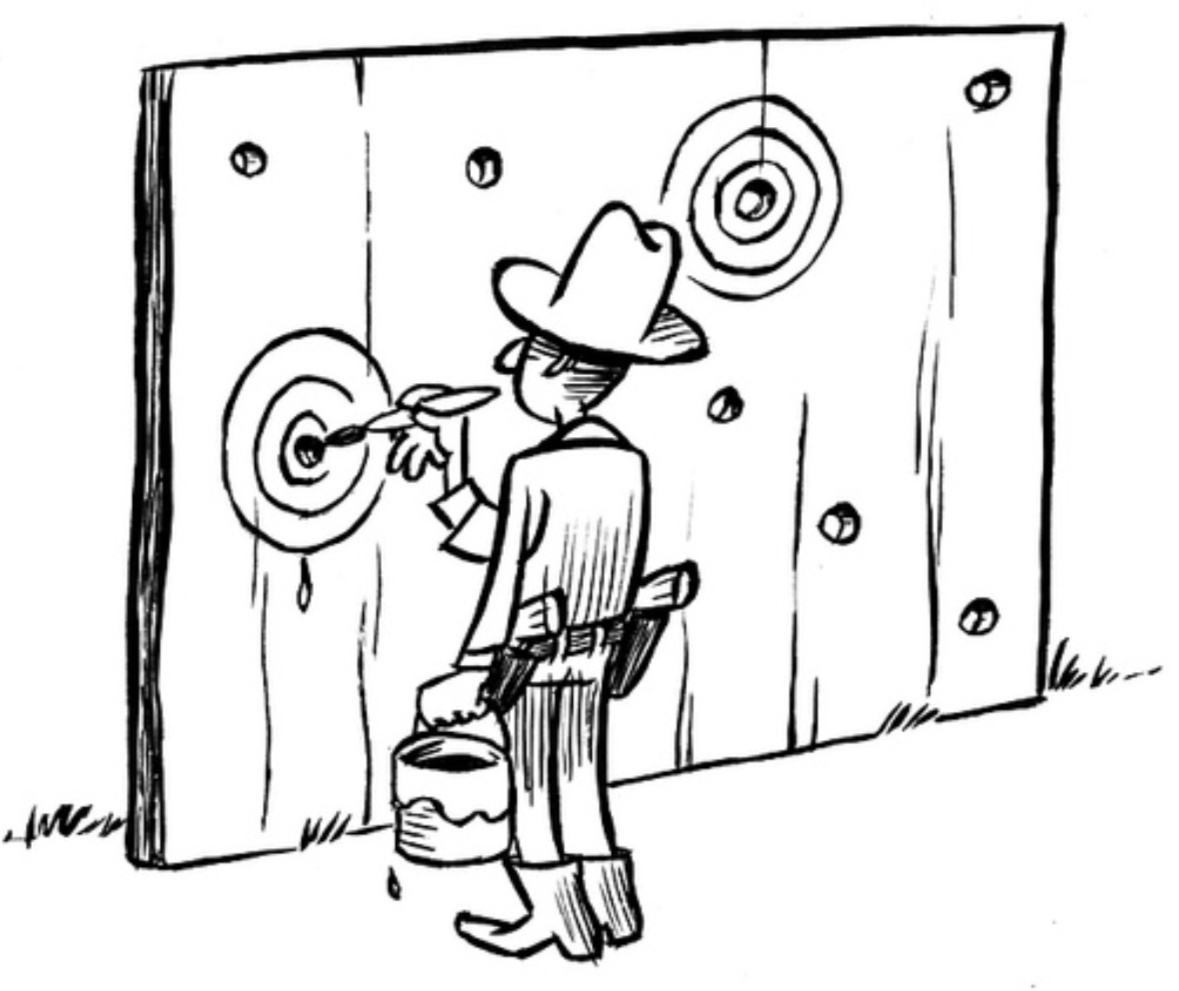
It’s not an artifact of our imagination that the house of cards is improbable. The house of cards is improbable in a way that resists both the Texas Sharpshooter Fallacy
and the Single Target Fallacy when there are multiple targets available:
The structure of the house of cards is improbable as a matter of principle, and so are things like DNA in the sense of the requisite orientation and positioning and connection of the monomers that form it. Vague appeals to some other chemical replicating machinery that doesn’t use DNA doesn’t make the emergence of DNA more probable!
Vague appeals to something not directly testable, not provable, not repeatable, not observable – that’s faith, not empiricism. Faith is fine as far as faith goes, but call a spade a spade, please, don’t call it empirical proof, call it FAITH.
Perry referred to RNA replicators as “life”. That’s the equivocation I was talking about – OOL researchers have redefined the problem so as to claim victory, when the problems isn’t making replicators subject to some sort of adaptive evolution. The problem is trying to show the improbable structure of cellular replicators isn’t really improbable – somewhat like trying to show the improbable structure of the house of cards above isn’t really improbable, that some mindless mechanism that obeys random distributions can put it together.
How improbable does something have to be before one accepts a miracle? Jon Perry said he would abandon OOL if there were another scientific research program. That’s reasonable, but if miracles aren’t repeatable, then how how will he, via a research program that searches for repeatable mechanisms ever reasonably infer that a miracle happened IF indeed a miracle was the cause of life? Miracles by definition aren’t repeatable on demand like laboratory chemistry. So Perry won’t get the “proof” of a miracle he is searching for. It is, on some level akin to the problem I posed to Tracie Harris regarding the blind individuals healed by prayer…
Whether life was a miracle is probably an undecidable proposition either way, and it’s something of a faith statement for each side of the argument. Same for the miracles that people may experience in their personal life. This raises the issue of the hiddeness of God and His selective distribution of miracles to some people and not others, (Prov 25:2, Matt 11:23, Matt 11:25, 1 Cor 1:26-29 etc.).
But, what Tan and Stadler’s book has done is shown, it’s premature at best to say OOL has progressed as a discipline in making naturalistic OOL of cellular life look solvable. Asphalt looks like the probable trajectory of biotic precursors, not life.